


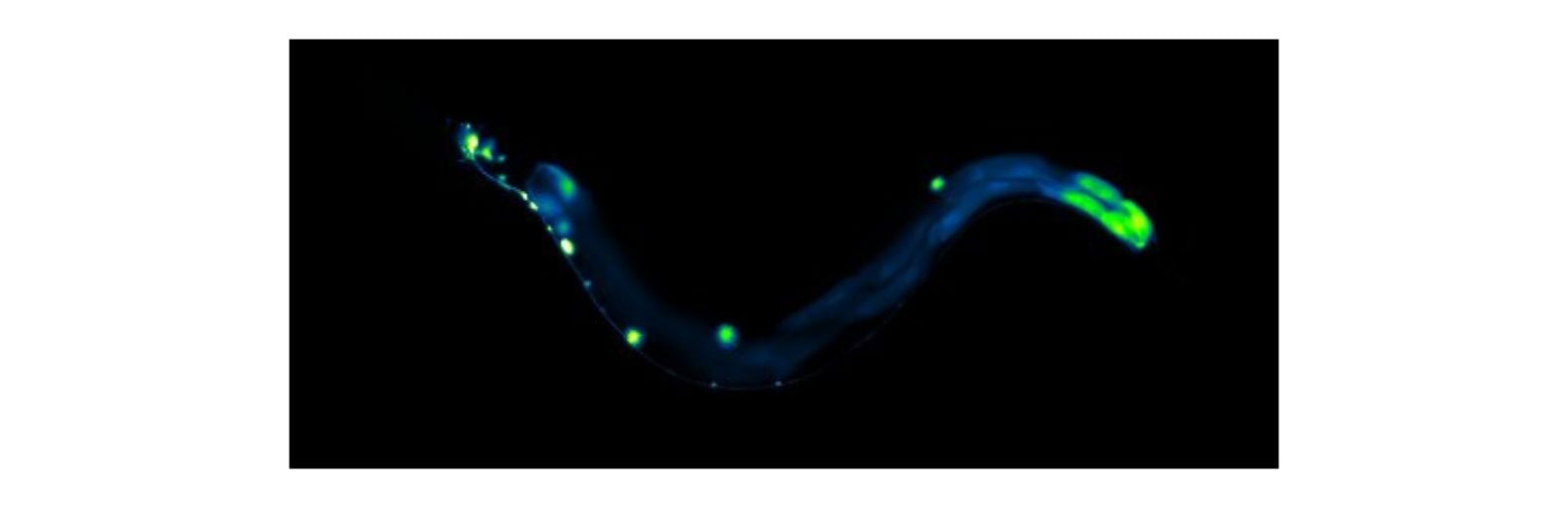
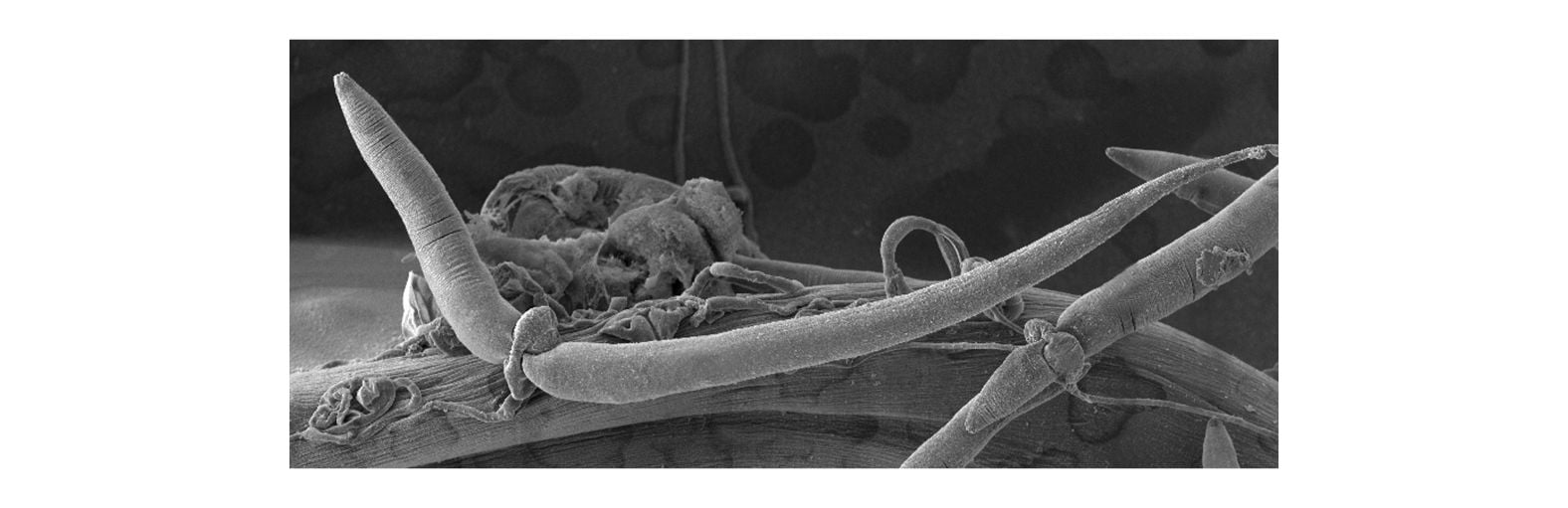
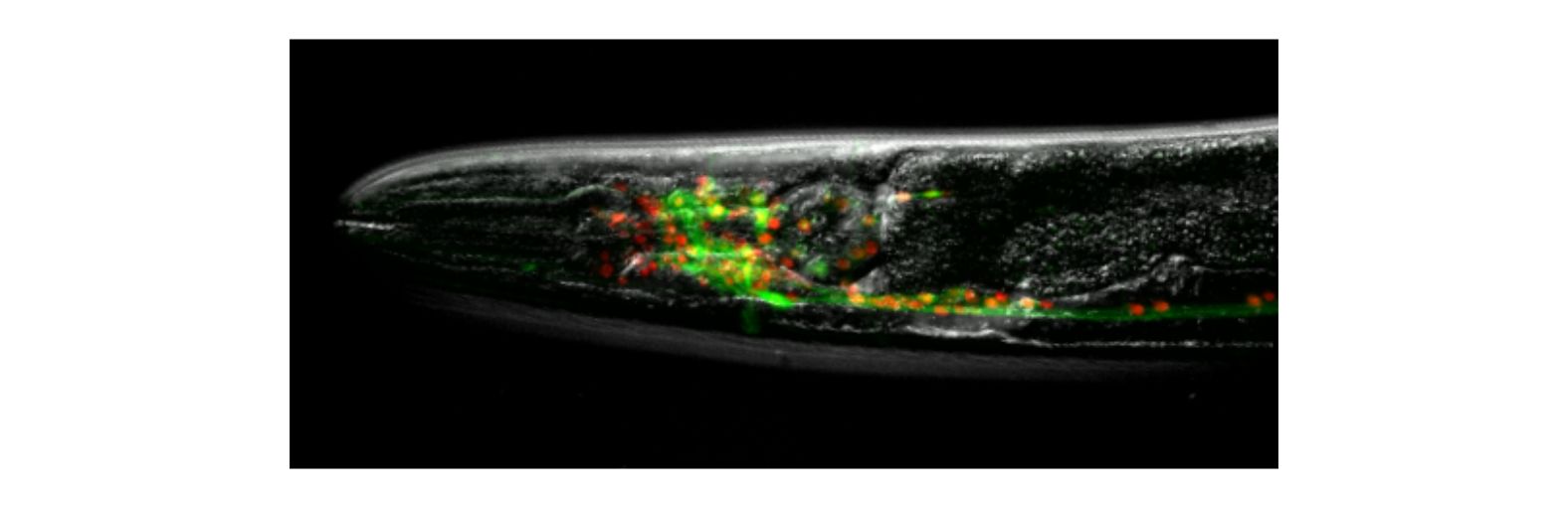
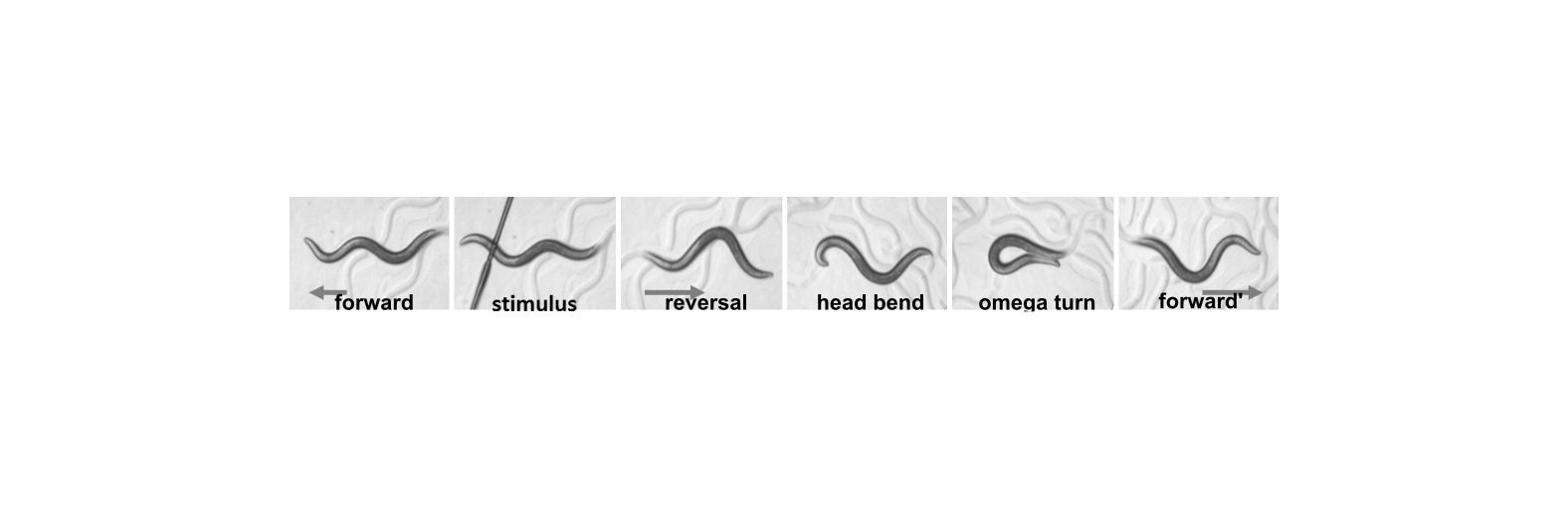


Our focus is to understand how the nervous system controls behavior and responds to changes in its environment.
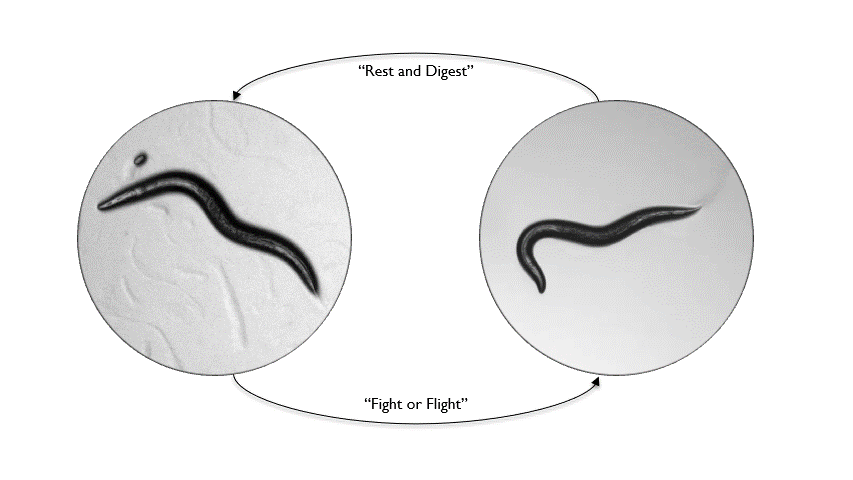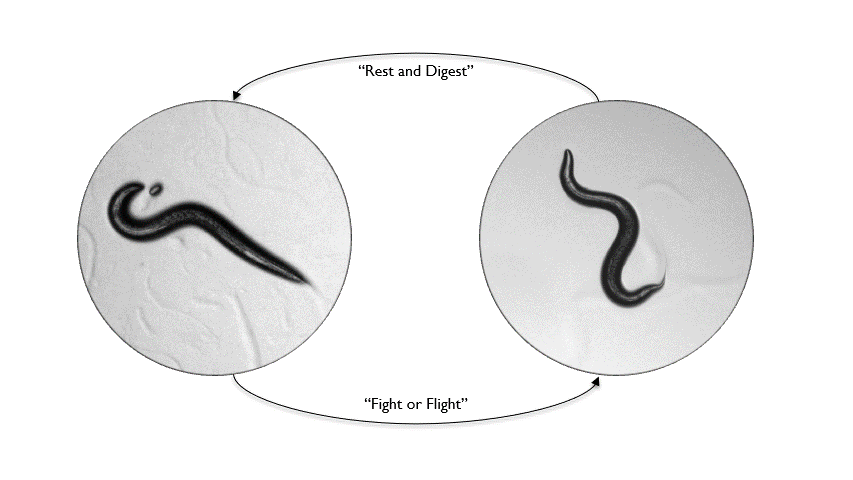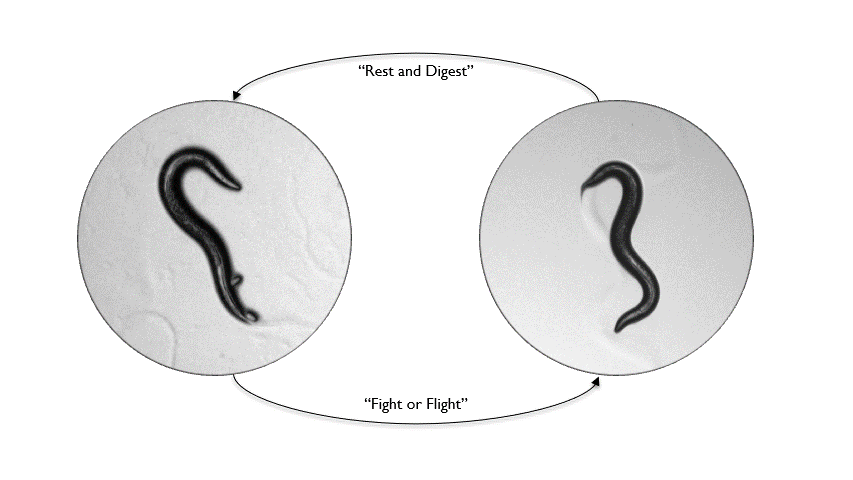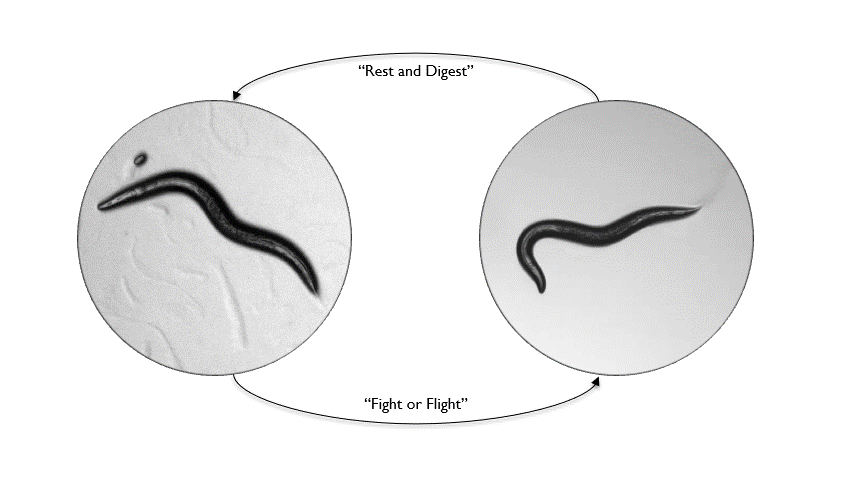
Animals inhabit complex and unpredictable environments where the availability of food and the presence of predators can change rapidly. To survive, they must adapt their behavior and physiological states flexibly. Monoamines, such as serotonin, dopamine, and norepinephrine, are neuromodulators that influence neural circuits over extended periods.Our research aims to understand how monoamine signaling shapes and modifies behavioral states in C. elegans. We employ genetics, automated behavioral analysis, and in vivo techniques to record and manipulate neuronal activity.
By examining foraging strategies under varying environmental conditions, such as food scarcity or predatory threats, we seek to elucidate the roles of individual monoamines in coordinating neural circuit activity, behavior, and physiology in a dynamic world.
~
How does the microbiome affect neuronal function and behavior?
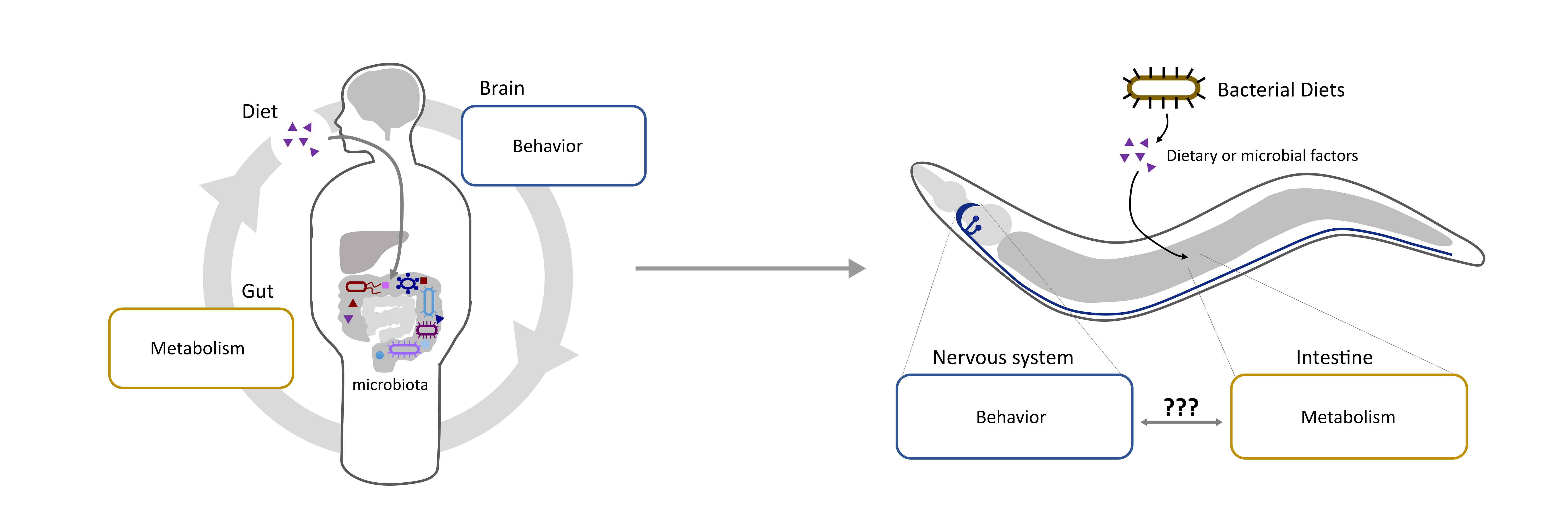
The human microbiome is composed of hundreds of different species of bacteria and can have a profound impact on diverse physiological processes, but unraveling the underlying mechanisms is a major challenge in humans due to the complexity of the nervous system and the diversity of microbial fauna present in the gut.
Using C. elegans as a model we can begin to address these questions in a meaningful way.
.